
| початок розділу
Виробничі, аматорські радіоаматорські Авіамодельний, ракетомодельного Корисні, цікаві |
хитрощі майстру
електроніка фізика технології винаходи |
таємниці космосу
таємниці Землі таємниці Океану хитрощі Карта розділу |
  |
| Використання матеріалів сайту дозволяється за умови посилання (для сайтів - гіперпосилання) | |||
Навігація: => |
На головну / Каталог патентів / В розділ каталогу / Назад / |
|
ВИНАХІД
Патент Російської Федерації RU2215407
![]()
СПОСІБ СТВОРЕННЯ ВИХІДНОГО МАТЕРІАЛУ ДЛЯ СЕЛЕКЦІЇ РОСЛИН
Ім'я винахідника: Зеленцов С.В .; Кочегура А.В.
Ім'я патентовласника: Державна наукова установа РСХН - Всеросійський науково-дослідний інститут олійних культур ім. В.С. Пустовійта
Адреса для листування: 350038, г.Краснодар, вул. Філатова, 17, ДНУ РАСГН ВНІІМК, патентна група
Дата початку дії патенту: 2002.01.03
Винахід призначений для використання в галузі сільського господарства. Спосіб включає переклад вихідних диплоїдних сортів на тетраплоїдний рівень колхіцінірованіем або іншим відомим способом. Виділяють реверсивні диплоїдні рослини за морфологічними та іншим господарсько цінними ознаками і використовують їх як вихідний матеріал. Отримують поліплоїди подвоєнням плоїдності вихідних сортів з будь-яким її початковим рівнем і вирощують їх протягом декількох поколінь. Одночасно з цим в кожному поколінні відбирають рекомбинированного поліплоїдні форми, що відрізняються від базового поліплоїдного морфотіпа. Відібрані форми розмножують, кожну окремо, протягом ряду поколінь, в кожному з яких виділяють форми з реверсивно кратнопоніженной плоїдності - реплоіди за морфологічними, біохімічними або іншими ознаками, рівень плоїдності яких визначають прямим підрахунком хромосом. Оцінюють реплоіди за комплексом господарсько цінних ознак, виділяючи форми, що відрізняються від рослин з вихідною плоїдності зміненими або новими кількісними або якісними ознаками. Винахід дозволяє збільшити поліморфізм кількісних і якісних ознак рослин, отримати нові ознаки, а й повернути втрачені в процесі філогенезу ознаки без використання мутагенів різної природи.
ОПИС ВИНАХОДИ
Винахід відноситься до сільського господарства і може бути використано в селекції рослин.
Відомий спосіб селекції рослин, що включає схрещування штучно зміненої форми і вихідного сорту, відбір цінних форм серед гібридів і їх розмноження, причому в якості штучно зміненої форми використовують міксоплоіди, які в схрещуванні беруть участь в якості материнського компонента. Необхідний міксоплоід, наприклад пшениці, отримують експериментальним замочуванням сухого насіння у водному розчині 5% -ного колхіцину протягом 24 год при 25 o С і висіву їх на постійне місце в поле без попередньої промивки у воді (А. с. СРСР 1292681, А 01 Н 1/04, публ. 28.02.87 р, Бюлл. 8).
Недоліком відомого способу є те, що в якості штучно зміненої форми для схрещування використовують міксоплоіди, що відрізняються змішаної плоїдності (вихідної, подвоєною і анеуплоїдних) генома. При цьому видимі міксоплоідние морфологічні зміни колоса, що належать материнському рослині не дозволяють передбачити рівень плоїдності макро- і мікрогамет в кожній квітці колоса пшениці або ячменю. Беруть участь в схрещуванні квітки можуть мати як вихідну, так і змінену плоїдність. При схрещуванні квітки з незміненою плоїдності на міксоплоідном колосі з вихідним нормальним сортом покоління F 1 буде повністю гомозиготною і аналогічною початковому сорту. При схрещуванні вихідного сорту, наприклад гексаплоїдної (2n = 6х = 42) м'якої пшениці або ярого ячменю (2n = 2х = 14), з квіткою подвоєною плоїдності (4n = 12х = 84 або 4n = 4x = 28 відповідно) на міксоплоідном колосі, будуть отримані зиготи проміжної плоидности (3n = 9х = 63 або 3n = 3х = 21 відповідно) з вираженими порушеннями кон'югації хромосом в профазі I мейозу I через некратні їх числа і подальшого несиметричного розбіжності хромосом в анафазе мейозу. Наслідком цього буде формування яскраво вираженою стерильності, гноблення, сублетальні або летальності гібридних рослин. Використання в схрещуванні анеуплоїдних квіток міксоплоідного колоса з числом хромосом більше або менше на 1 або 2 хромосоми щодо базового генома призведе до утворення уніваленти в мейозі через наявність непарних хромосом і в багатьох випадках до наступної повної стерильності або летальності рослин. В цілому, використання міксоплоідов для схрещування з вихідними сортами через високу частку утворюються форм з проміжної (3n) плоїдності або Анеуплоїдія і пов'язаної з цим високою летальністю або стерильністю не дозволяє виявляти і закріплювати в потомстві велику кількість корисних рекомбінацій і не забезпечує збільшення поліморфізму кількісних і якісних ознак у сортів і видів рослин.
Відомий спосіб селекції ячменю, що включає переклад диплоїдних сортів на тетраплоїдний рівень, отримання тетраплоідних форм, проведення штучної деполіплоідізаціі аутотетраплоідов в поколінні не нижче З 5 шляхом впливу на вирощувані в судинах проростки температурою 5-10 o С і 7-8-годинним освітленням і подальше виділення диплоїдних реверсантов - за розмірами клітин первинних корінців, які повинні бути близькі до розмірів клітин тетраплоідних форм (А.с. СРСР 1591885, А 01 Н 1/04, публ. 1990 року).
Недоліком відомого способу є те, що для деполіплоідізаціі використовуються тетраплоїди не раніше ніж 5-го покоління (С 5), при цьому в відборі не беруть участь більш ранні покоління автотетраплоідов (C 2 -C 4) з найбільшою частотою рекомбінації ознак. При використанні даного способу диплоїдним реверсантам передаються цитоплазматичних детермініруемие ознаки і не враховується більшість господарсько цінних ознак, контрольованих ядерними хромосомами. Виявлення диплоїдних реверсантов з ознаками тетраплоїдів тільки методом визначення розмірів клітин корінців без прямого підрахунку плоїдності сприяє втраті частини диплоїдних реверсантов, а й відбору великої кількості стабілізованих тетраплоїдів з більшими у порівнянні з диплоїдним сортом морфологічними елементами рослини, як це наведено в описі відомого способу, що відрізняються , як правило, пухкої соматичної тканиною і зниженим внутрішньоклітинним осмотичним тиском. Штучна деполіплоідізація тетраплоїдів в судинах істотно знижує ефективність способу за рахунок обмеженого числа рослин, що визначається числом судин і розмірами споруд штучного клімату. Знижена, 12-15 o С, температура при деполіплоідізаціі вимагає додаткових енерговитрат і устаткування на підтримку необхідних температурних режимів. В цілому, застосування відомого способу істотно обмежує виявлення можливого поліморфізму кількісних і якісних ознак через те, що корисні рекомбінації, що утворюються в більш ранніх, ніж З 5, поколіннях, не виділяються і не використовуються, а й з-за відбору обмеженого обсягу форм рослин невстановленої плоидности (в т.ч. фертильних стабілізованих тетраплоїдів), іменованих у відомому способі диплоїдними реверсантамі, що визначається технічними умовами реверсії плоїдності в контрольованих умовах середовища.
Відомий і спосіб створення вихідного матеріалу для селекції зернових і круп'яних культур, що включає переклад диплоїдних сортів на тетраплоїдний рівень колхіцінірованіем або іншим відомим способом, обробку насіння тетраплоїдів ![]() -випромінюванням або розчином нітрозоетілмочевіни в дозі, критичної для диплоидной форми, вирощування поколінь M 1 і М 2 у вигляді популяцій, виділення в поколінні М 2 реверсивних диплоїдних рослин (за такими ознаками: велика висота рослин, раннє колосіння, висока озерненность колоса), використання насіння з відібраних рослин для закладки ліній (А.с. СРСР 1519596, А 01 Н 1/04, публ. 1989 г.).
-випромінюванням або розчином нітрозоетілмочевіни в дозі, критичної для диплоидной форми, вирощування поколінь M 1 і М 2 у вигляді популяцій, виділення в поколінні М 2 реверсивних диплоїдних рослин (за такими ознаками: велика висота рослин, раннє колосіння, висока озерненность колоса), використання насіння з відібраних рослин для закладки ліній (А.с. СРСР 1519596, А 01 Н 1/04, публ. 1989 г.).
Недоліком відомого способу, взятого нами як прототип, є те, що метою даного винаходу є отримання мутантів реверсивно диплоидного походження. Використання фізичних або хімічних мутагенів в критичних дозах (LD 50) призводить до модифікації господарсько цінних ознак за рахунок порушення функціональності окремих локусів, що контролюють їх реалізацію. При цьому в геномі рослини збільшується частка морфологічно непомітних мутацій і мікромутацій, надають адитивно-негативну дію на життєздатність організму в цілому. Як правило, всі ознаки, отримані шляхом мутагенезу, в тому числі і при використанні відомого методу, взятого нами як прототип, рецесивні і вимагають підтримки організму в гомозиготному стані. Частота народження реверсивних диплоидов при використанні ![]() -випромінювання має яскраво виражену сортову залежність, а їх різноманітність визначається тільки кількісними ознаками, контролюються полімерно (велика висота рослин, раннє колосіння, висока озерненность колоса). Використовуються в прототипі прийоми для збільшення частоти появи реверсивних диплоидов істотно обмежують можливості для відбору господарсько цінних диплоїдних реверсантов по модифікованим кількісним і якісним ознаками, оскільки відбір реверсантов ведеться тільки після обробки мутагенами в критичних дозах (тобто при мутагенної летальності не менше 50% особин ). При цьому частина корисних генних рекомбінацій, що утворилися на Поліплоїдний рівні або при реверсії на диплоїдний рівень, буде загублена через утворення несумісних з життям організму мутацій, викликаних застосуванням критичних доз мутагенів, а більшість новоутворених ознак, в т.ч. і спочатку домінантних у вихідного сорту або тетраплоїда, придбають рецессивность через часткове пошкодження і порушення функціональності мутованих локусів. В цілому, відомий спосіб, обраний нами як прототип, істотно обмежує отримання, виявлення і подальше збереження максимально можливого поліморфізму за кількісними та якісними ознаками рослин.
-випромінювання має яскраво виражену сортову залежність, а їх різноманітність визначається тільки кількісними ознаками, контролюються полімерно (велика висота рослин, раннє колосіння, висока озерненность колоса). Використовуються в прототипі прийоми для збільшення частоти появи реверсивних диплоидов істотно обмежують можливості для відбору господарсько цінних диплоїдних реверсантов по модифікованим кількісним і якісним ознаками, оскільки відбір реверсантов ведеться тільки після обробки мутагенами в критичних дозах (тобто при мутагенної летальності не менше 50% особин ). При цьому частина корисних генних рекомбінацій, що утворилися на Поліплоїдний рівні або при реверсії на диплоїдний рівень, буде загублена через утворення несумісних з життям організму мутацій, викликаних застосуванням критичних доз мутагенів, а більшість новоутворених ознак, в т.ч. і спочатку домінантних у вихідного сорту або тетраплоїда, придбають рецессивность через часткове пошкодження і порушення функціональності мутованих локусів. В цілому, відомий спосіб, обраний нами як прототип, істотно обмежує отримання, виявлення і подальше збереження максимально можливого поліморфізму за кількісними та якісними ознаками рослин.
Заявляється винаходом вирішується завдання підвищення ефективності способу за рахунок розширення поліморфізму як домінантних, так і рецесивних алелей якісних і кількісних ознак при отриманні реверсантов (реплоіди) (вперше введений нами термін) у будь-яких видів вищих рослин, які не досягли критичного рівня плоїдності.
Мета винаходу - збільшення поліморфізму кількісних і якісних ознак рослин, а й отримання нових ознак або повернення втрачених в процесі філогенезу виду ознак, без використання мутагенів різної природи, шляхом поліплоїдизації рослин, подальшого відбору рекомбінантних полиплоидов і виділення серед них за рахунок спонтанної редукції плоїдності гамет форм з поворотної кратно-зниженою плоїдності - реплоіди.
Технічний результат досягається тим, що у відомому способі, що включає переклад вихідних диплоїдних сортів на тетраплоїдний рівень колхіцінірованіем або іншим відомим способом, виділення реверсивних диплоїдних рослин за морфологічними та іншим господарсько цінними ознаками та використання їх в якості вихідного матеріалу, відповідно до винаходу отримують поліплоїди подвоєнням плоїдності вихідних сортів з будь-яким її початковим рівнем, вирощують поліплоїди протягом декількох поколінь і одночасно з цим відбирають рекомбинированного поліплоїдні форми, що відрізняються від базового поліплоїдного морфотіпа, відібрані форми розмножують, кожну окремо, протягом ряду поколінь, в кожному з яких виділяють форми з реверсивно кратно -зниження плоїдності (реплоіди), рівень плоїдності яких визначають за морфологічними ознаками і прямим підрахунком хромосом, оцінюють реплоіди за комплексом господарсько цінних ознак, виділяючи форми, що відрізняються від рослин з вихідною плоїдності зміненими або новими якісними і кількісними ознаками.
Порівняльний аналіз заявляється технічного рішення з прототипом дозволяє зробити висновок, що заявляється спосіб відрізняється від відомого наявністю нового прийому: виділенням рекомбинированного полиплоидов з внутрісортовой популяції полиплоидов, отриманих подвоєнням плоїдності вихідних сортів і вирощуваних протягом декількох поколінь, а й порядком виконання відомого прийому "виділення реверсивних форм ": форми з реверсивно кратно-зниженою плоїдності (реплоіди) виділяють з рекомбинированного поліплоїдних форм при їх розмноженні протягом ряду поколінь. Таким чином, заявляється спосіб відповідає критерію патентоспроможності "НОВИЗНА".
Досліджуючи рівень техніки в процесі проведення патентного пошуку за всіма видами відомостей, загальнодоступних у пресі, ми виявили, що спосіб створення вихідного матеріалу для селекції рослин, що включає заявлену нами сукупність ознак, для фахівця в галузі селекції явно не випливає з відомого на сьогодні існуючого рівня техніки. Нашими дослідженнями протягом ряду років виявлено, що в природі з певною частотою постійно здійснюється спонтанна полиплоидизация вищих рослин. Поліплоїдизація призводить до утворення полівалентов в профазі I мейозу I. З цим пов'язані різні порушення кон'югації і кроссинговера гомологічниххромосом, що викликають їх структурні зміни. Нерегулярне розподіл дочірніх хромосом в мейозі у "ранніх" автополіплоїдов призводить до редукції плоїдності гамет і утворення Анеуплоїдія і зворотних диплоидов (реплоіди). У "ранніх" автополіплоїдов при мультівалентной кон'югації і кроссинговере можуть відбуватися асиметричні обміни гомологічними ділянками хромосом. Структурні зміни хромосом можуть призводити до кількісних і якісних змін існуючих або до утворення нових ознак. Реплоідние форми утворюються на основі складових поліплоїдний геном диплоїдних геномів, включаючи рекомбинированного набори хромосом. При цьому утворилися реплоіди можуть бути гомологічними вихідної диплоїдної формі або відрізнятися від неї, зберігаючи виникли на Поліплоїдний рівні нові ознаки або купуючи їх при мультівалентной кон'югації і нерівному кроссинговере, що передує редукції плоїдності. Реплоіди, що відрізняються зміненими якісними і кількісними ознаками, а й новими ознаками, в т.ч. загубленими в процесі філогенезу виду, можуть використовуватися в якості вихідного матеріалу при селекції рослин. На підставі вищевикладеного можна зробити висновок про відповідність заявляється технічного рішення критерію патентоспроможності "ВИНАХІДНИЦЬКИЙ РІВЕНЬ".
Заявляється технічне рішення відповідає і критерієм патентоспроможності "промислова придатність", тому що воно може бути використано в сільському господарстві, і, крім того, в описі винаходу будуть представлені засоби і методи, за допомогою яких можливе здійснення технічного рішення в тому вигляді, як воно охарактеризовано в незалежному пункті формули винаходу.
Спосіб здійснюють наступним чином. Починають з того, що беруть сорти або форми рослин з вихідною плоїдності (наприклад, діплоїден) і переводять їх на поліплоїдний (наприклад, тетраплоїдний) рівень колхіцінірованіем або іншим відомим способом. У першому Поліплоїдний поколінні (C 1) виділяють поліплоїди за характерними морфологічними ознаками, властивим полиплоидам даного виду, і прямим підрахунком хромосом. Виділені поліплоїди вирощують (в польових умовах або в умовах штучного клімату) протягом декількох поколінь. У кожному Поліплоїдний поколінні відбирають рекомбинированного поліплоїдні форми, явно відрізняються від базового поліплоїдного морфотіпа (по будь-яким кількісним і якісним ознаками), що свідчить про структурні зміни хромосом одного або більше складових поліплоїдії диплоїдних геномів. Виділені рекомбинированного поліплоїдні форми вирощують протягом декількох поколінь окремими популяціями, виділяючи в кожному поколінні реплоіди зі зворотно кратнопоніженной плоїдності за морфологічними ознаками і прямим підрахунком хромосом. Оцінюють реплоіди за комплексом господарсько цінних ознак, виділяючи форми, що відрізняються від рослин з вихідною плоїдності зміненими або новими якісними і кількісними ознаками. Всі виділені реплоіди, що відрізняються зміненими, поліпшеними або новими морфологічними, біохімічними або іншими ознаками, використовують в якості вихідного матеріалу в селекційних чи інших цілях.
Приклад здійснення способу
В якості вихідних форм були взяті сорти сої Фора і Віла з 2n = 40. Їх насіння висіяли в польових умовах і в фазу 1-го трійчастого листа рослин перевели на поліплоїдний рівень шляхом обробки апікальних меристем водними розчинами колхіцину в концентрації 0,05%. Утворені нижче зони колхіцінірованія диплоїдні пагони видаляли у міру їх появи. Колхіцінірованние пагони доводили до дозрівання і на них прибирали насіння З 1.
Отримане насіння З 1 маркували і індивідуально пророщували в чашках Петрі до отримання корінців довжиною 2-5 см, відрізали кінчик корінця довжиною 0,5 см і цитологічних досліджували стандартними методиками. Проростки, у яких була встановлена подвоєна плоїдність (4n = 80), висівали в грунт, формуючи по кожному сорту Поліплоїдний популяцію C 1.
Починаючи з покоління C 1 у внутрісортових популяціях полиплоидов сої щорічно виділяли рекомбинированного поліплоїдні форми, явно відрізняються від базового поліплоїдного морфотіпа по швидкості і тривалості етапів органогенезу, висоті рослин, характеру розгалуження, формі і числу листочків справжніх листків, довжині черешка, забарвленням опушення, гіпокотиля, віночка квіток, оболонки і рубчика насіння (табл. 1). Оскільки соя є строгим самоопилітель, і крім цього, гібридизація між діплоїден і тетраплоїдом у сої виключена через нежиттєздатність тріплоідний зигот, поява змінених, генетично детермінованих ознак у внутрісортових поліплоїдних популяціях свідчить про поліплоїдні рекомбінації геному без залучення генетичного матеріалу ззовні.
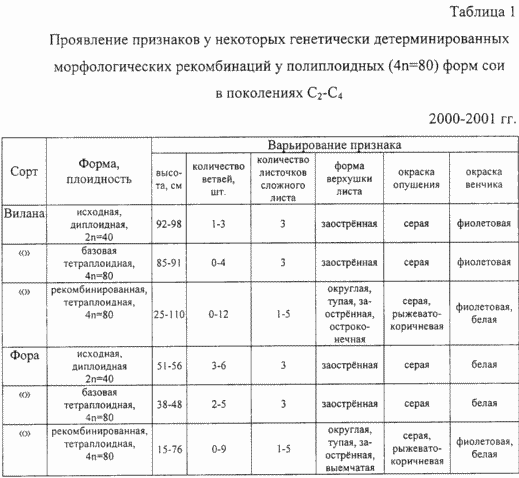
Аналіз табл. 1 свідчить про наявність поліморфізму по виявленим рекомбінованим ознаками поліплоїдних (4n = 80) форм сої. Так, при рекомбінації ознаки, контролюючого формування трійчастого складного листа диплоїдного сорту і базової поліплоїдні форми у поліплоїдних рекомбінантов, кількість складових складний лист листових пластинок варіював від п'яти листочків (дуже рідко зустрічається форми) до однолісточковой форми, повністю відсутньою в природному генофонді виду. Виявлено поліморфізм у рекомбінантов за формою верхівки листа, що варіює від гострої до тупий і виїмчастою. Забарвлення опушения стебел, що має сіре забарвлення у обох диплоїдних сортів і їх базових поліплоїдних форм, у окремих полиплоидов набувала рудувато-коричневе забарвлення, відтворену в наступних поколіннях. У деяких рекомбінантних полиплоидов белоцветкових сорти Фора була виявлена фіолетове забарвлення віночка, а у сорту Віла, що має фіолетові квітки, - були виділені белоцветкових поліплоїди. У рекомбінантних полиплоидов обох сортів помітно збільшився діапазон мінливості по висоті рослин від карликових до високорослих форм, а й за кількістю гілок.
У потомстві рекомбинированного полиплоидов щороку виділяли реплоіди по візуально помітним морфологічними ознаками, властивим сої з вихідної базової плоїдності: пропорціям листових пластинок, розмірами пилкових зерен і кількістю на їх поверхні мікропор, формі, характеру поверхні і розмірами бобів і насіння і перевіряли рівень плоїдності реплоіди прямим підрахунком хромосом.
Виділені реплоіди сої порівнювали за комплексом якісних і кількісних ознак в порівнянні з вихідним диплоїдним сортом, виділяли форми з поліпшеними господарсько цінними ознаками і використовували їх як вихідний матеріал для селекції сої (табл. 2-3).
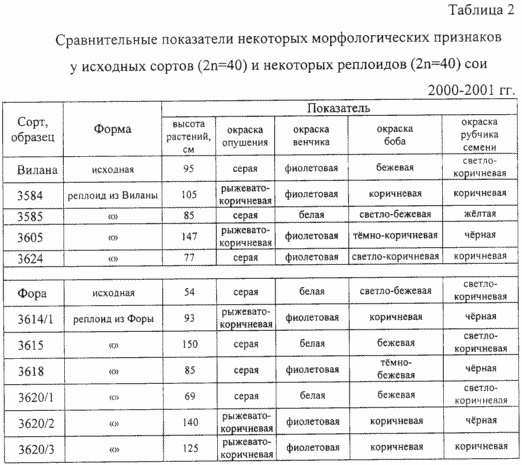
Аналіз табл. 2 свідчить про наявність морфологічних рекомбінації по ряду ознак у реплоіди, виділених з рекомбинированного полиплоидов. Так, висота реплоідних рослин варіювала від 85 до 147 см у сорту Віла і від 69 до 150 см у сорту Фора, в окремих випадках в 3 рази перевищуючи висоту вихідного сорту. Були виділені реплоіди, що відрізняються від вихідних сортів за кольором забарвлення опушення стебел і віночка квіток. Виявлено поліморфізм за такими якісними ознаками, як забарвлення стулок бобів, що варіює від світло-бежевою до коричневої і темно-коричневої, і забарвлення рубчика насіння, що варіює від жовтої до чорної.
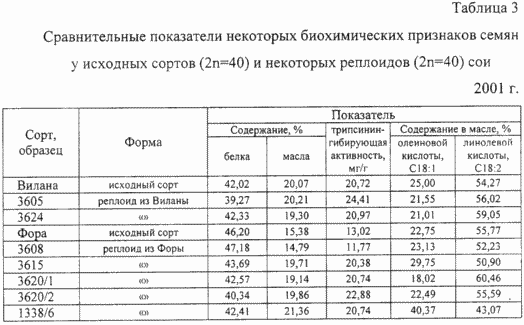
Виділені реплоіди відрізнялися від вихідних сортів і і за біохімічними, полімерно спадкоємною ознаками (табл.3). Так, вміст білка в насінні реплоіди відрізнялося від рівня вихідних сортів: на 2,75 абсолютних% у сорту Віла і на 5,86 абсолютних% у сорту Фора. У сорту Фора відзначений поліморфізм за змістом масла в насінні, що варіює від 14,79 до 19,86%. Поліплоїдні рекомбінації піддався і показник тріпсінінгібірующей активності білка, в окремих випадках збільшуючись на 3,69 мг / г у реплоіди сорти Віла, а у реплоіди нізкоінгібіторного сорти Фора - варіюючи від 11,77 до 22,88 мг / г при вихідному рівні 13,02 мг / г. Аналіз жирно-кислотного складу олії у реплоіди сої і показав наявність генетичних рекомбінацій рівня біохімічного синтезу окремих жирних кислот. У реплоіди сорти Віла відзначено зниження на 4 абсолютних% змісту олеїнової кислоти з одночасним збільшенням на 2-5 абсолютних% змісту лінолевої кислоти. Жирно-кислотний склад масла у реплоіди низькоолійне сорти Фора відрізнявся більш широким поліморфізмом і за змістом олеїнової кислоти варіював від 18,02 до 40,37% при вихідному рівні 22,75%, а за змістом лінолевої кислоти - від 43,07 до 60, 46% при вихідному вмісті 55,77%.
Жирно-кислотний склад соєвого масла зі зниженим, менше 20%, вмістом олеїнової кислоти і підвищеним, до 60%, вмістом лінолевої кислоти близький до балансу жирних кислот в олії дикорослих предків сої.
Жирно-кислотний склад масла реплоіди з сорту Фора 1338/6 з вмістом олеїнової кислоти 40,37%, подвоєним в порівнянні з вихідним сортом, і зі зниженим до 43,07% вмістом лінолевої кислоти є аналогом кунжутного масла і практично не невідомий в природному генофонді сої і, отже, є прикладом нового ознаки, створеного заявляється способом.
Аналогічним способом можна отримувати реплоіди зі зміненими або новими кількісними і якісними ознаками у будь-якого виду вищих рослин.
Отримання полиплоидов подвоєнням плоїдності вихідних сортів з будь-яким її початковим рівнем забезпечує зміна існуючих або поява нових ознак за рахунок порушень кон'югації і кроссинговера мультівалентних комплексів хромосом в мейозі, що супроводжуються структурними змінами окремих ділянок (локусів).
Вирощування полиплоидов протягом декількох поколінь і одночасний з цим відбір в кожному поколінні рекомбинированного поліплоїдних форм, що відрізняються від базового поліплоїдного морфотіпа, дозволяє виявити і зберегти все візуально помітні морфологічні рекомбінації на Поліплоїдний рівні, т. Е. Розширити, таким чином, існуючий поліморфізм ознак у полиплоидов за кількісними та якісними ознаками без використання мутагенів різної природи і сформувати популяцію полиплоидних рекомбінантов для подальшого виділення реплоіди.
Розмноження відібраних рекомбінантних поліплоїдних форм, кожної окремо, протягом ряду поколінь і виділення реплоідних форм забезпечує підвищення частоти виникнення реплоіди з ознаками генетично детермінованих рекомбінації, що утворилися на Поліплоїдний рівні або в процесі деполіплоідізаціі при редукції плоїдності гамет, тобто дозволяє розширити поліморфізм кількісних і якісних ознак на реплоідном рівні без використання мутагенів різної природи.
Визначення рівня плоїдності реплоіди прямим підрахунком хромосом забезпечує достовірну ідентифікацію справжніх реплоіди з ознаками генетичної рекомбінації за кількістю хромосом, виключаючи віднесення до цієї категорії стабілізованих полиплоидов і Анеуплоїдія, і морфологічно відрізняються від вихідних сортів або видів рослин і їх базових поліплоїдних морфотипов, тобто забезпечує збільшення поліморфізму стабільно успадкованих кількісних і якісних ознак, а й нових або загублених в процесі філогенезу виду ознак, у справжніх реплоіди.
Оцінка реплоіди за комплексом господарсько цінних ознак з виділенням форм, що відрізняються від рослин з вихідною плоїдності зміненими або новими якісними і кількісними ознаками, дозволяє виявити і використовувати в селекції рослин всі позитивні генетично детерміновані рекомбінації, тобто розширити, таким чином, існуючий в природному генофонді виду рослин поліморфізм ознак.
Таким чином, заявляється винаходом забезпечується досягнення поставленої мети і рішення поставленого завдання.
ФОРМУЛА ВИНАХОДУ
1. Спосіб створення вихідного матеріалу для селекції рослин, що включає переклад вихідних диплоїдних сортів на тетраплоїдний рівень колхіцінірованіем або іншим відомим способом, виділення реверсивних диплоїдних рослин за морфологічними та іншим господарсько цінними ознаками та використання їх в якості вихідного матеріалу, який відрізняється тим, що отримують поліплоїди подвоєнням плоидности вихідних сортів з будь-яким її початковим рівнем, вирощують поліплоїди протягом декількох поколінь і одночасно з цим в кожному поколінні відбирають рекомбинированного поліплоїдні форми, що відрізняються від базового поліплоїдного морфотіпа, відібрані форми розмножують, кожну окремо, протягом ряду поколінь, в кожному з яких виділяють форми з реверсивно кратнопоніженной плоїдності - реплоіди, рівень плоїдності яких визначають прямим підрахунком хромосом, оцінюють реплоіди за комплексом господарсько цінних ознак, виділяючи форми, що відрізняються від рослин з вихідною плоїдності зміненими або новими кількісними або якісними ознаками.
2. Спосіб за п. 1, який відрізняється тим, що реплоіди виділяють по морфологічним, біохімічним або іншими ознаками.
Версія для друку
Дата публікації 11.03.2007гг
![]()

Коментарі
Коментуючи, пам'ятайте про те, що зміст і тон Вашого повідомлення можуть зачіпати почуття реальних людей, проявляйте повагу та толерантність до своїх співрозмовників навіть у тому випадку, якщо Ви не поділяєте їхню думку, Ваша поведінка за умов свободи висловлювань та анонімності, наданих інтернетом, змінює не тільки віртуальний, але й реальний світ. Всі коменти приховані з індексу, спам контролюється.